

- 首 页
- 关于我们
- 环境丰容
- 笼器具
- 实验器械
- 仪器设备
- 病理产品
- 试剂耗材
- 实验服务
- 科技学苑












































在毒理学研究中,了解所用非人灵长类动物的比较生理学,有助于毒理学家:1)更准确地判断毒理学发现对人类的相关性;2)针对合适的发育阶段设计实验;3)认识到动物模型在预测人类毒性方面的局限性;4)在对照组样本量较小时更合理地评估毒性变化;5)提高对可能无意中影响研究终点的混杂因素的认识。
其中,恒河猴(Macaca mulatta)是研究最为深入的非人灵长类动物,已有超过150年的科学研究历史。然而,1978年印度实施的贸易限制禁止了恒河猴出口,导致研究更加依赖实验室现有种群以及来自中国的出口。因此,食蟹猴(Macaca fascicularis,又称食蟹猕猴、长尾猕猴)在药物测试中的使用逐渐增多。当然,不仅因为上述供应变化,也因其体型略小、性格更温顺,且具有全年可繁殖(非季节性繁殖)的特点。
恒河猴和食蟹猴均分布于广阔的地理区域,已发现不同亚群在生物学特征上存在细微差异,可能影响实验结果。例如,源自毛里求斯的食蟹猴是一个隔离的外来种群,常用于生物医学研究,其血液学参数就与来自菲律宾和越南的食蟹猴有所不同。
此外,作为新世界猴的普通狨猴(Callithrix jacchus)在生理上与人类差异较大,但由于体型小、在人工繁育条件下每年可繁殖两次,也成为大型灵长类动物的一种有吸引力的替代选择。
遗传学对比
物种在形态和功能上的差异,被认为主要由细胞核和线粒体DNA所编码的遗传信息以及有限的表观遗传修饰所决定。早期研究通过显微镜观察染色体的条带模式和数量来评估物种间的遗传相似性。人类拥有46条二倍体染色体(包括2条性染色体),而类人猿(如黑猩猩等)有48条,长臂猿有44条,猕猴(食蟹猴、恒河猴)和狨猴的染色体数目则见于下表。
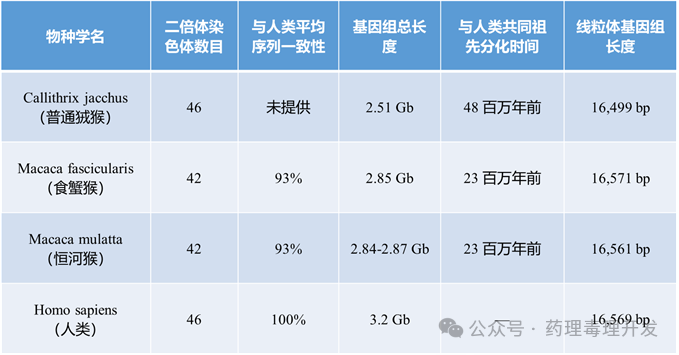
利用染色体条带技术对恒河猴和食蟹猴进行比较,发现两者染色体几乎完全同源,仅可能在5号染色体长臂上存在一个臂内倒位。线粒体基因分析表明,食蟹猴大约在150万至300万年前从恒河猴及其他猕猴中分化出来。但全基因组测序却发现,中国恒河猴的DNA(可能通过雄性个体迁入并与食蟹猴杂交)已渗入部分食蟹猴群体,从而增加了后者的遗传多样性。事实上,实验室中已成功培育出可存活的恒河猴–食蟹猴杂交后代。这种地理隔离程度及潜在的种间杂交现象,被认为是造成不同地区食蟹猴群体遗传多样性差异的原因。
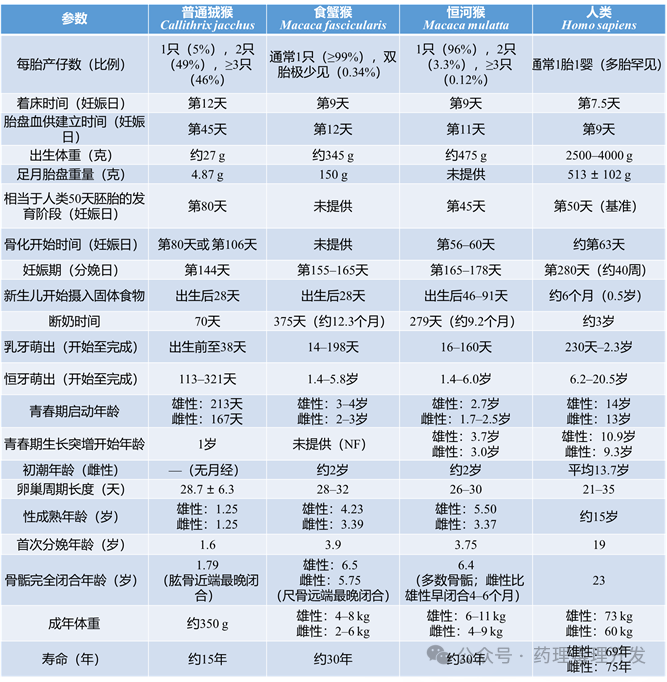
生长与发育对比
下表详细对比了狨猴(marmoset)、猕猴(macaques)和人类的关键发育里程碑。
(一)促性腺激素释放的激素调控
猕猴与人类高度相似的神经内分泌调控机制
猕猴的卵巢周期在神经内分泌调控方面与人类极为相似:下丘脑中基底部和弓状核的神经元合成并释放促性腺激素释放激素(GnRH)。GnRH通过垂体门脉系统作用于垂体前叶,刺激其释放两种促性腺激素:卵泡刺激素(FSH)和黄体生成素(LH)。GnRH是一种十肽,存在两种形式:GnRH-I和GnRH-II,分别由脑内不同神经元群体表达。两者均可通过GnRH受体-1刺激FSH和LH的释放。
目前认为:GnRH-I神经元负责卵巢周期大部分时间内的脉冲式FSH/LH释放;GnRH-II神经元则主要介导排卵时的LH峰。
2.狨猴(新世界猴)的独特性
普通狨猴等新世界猴具有一种第二型GnRH受体(GnRH-II受体),能特异性地响应GnRH-II配体。激活该受体似乎可促进性行为。值得注意的是:人类及其他旧世界猴(如猕猴)不表达GnRH-II受体。
3.LH受体缺失与CG的代偿机制
狨猴和其他新世界猴的LH受体(LH-R)在第10外显子存在缺失突变,导致其对LH不敏感,但仍能响应绒毛膜促性腺激素(CG)。作为补偿,这些动物不仅在胎盘、还在垂体中表达CG,从而在功能上以CG替代LH。CG的释放虽受GnRH调控,但其调控机制不完全等同于LH。
4.Kisspeptin的核心调控作用
来自垂体和性腺的分子信号通过上游两类神经元调控GnRH释放,而Kisspeptin(Kiss1)是关键的神经肽信使。
室周前腹侧核(AVPV)神经元:对雌性体内性激素敏感,介导正反馈(如排卵前雌激素升高→Kiss1↑→GnRH↑)。
弓状核(ARC)神经元:在所有物种中均对性激素敏感,介导负反馈(抑制Kiss1和GnRH释放)。
此外,室周核和前背侧视前核也表达Kiss1。Kiss1通过其受体GPR54激活GnRH神经元。Kiss1不仅在中枢神经系统表达,还存在于胎盘、睾丸、胰腺、肝脏和肠道中,其表达受瘦素(leptin)和胃饥饿素(ghrelin)调节,可能构成机体能量状态与生殖功能启动/维持之间的桥梁。
5.其他调控因素
雌激素还可通过快速调节GnRH神经元上的离子通道,直接影响其兴奋性和活动。多种神经递质系统(如谷氨酸、GABA、多巴胺等)也被证实参与GnRH释放的调控。
(二)卵巢周期(The Ovarian Cycle)
周期长度与阶段
食蟹猴:平均周期为30.4±4.7天。分为三个阶段,即卵泡期12–14天,围排卵期约3天,黄体期14–16天(若未妊娠则退化)
狨猴:总周期约28.7天。卵泡期较短(约8天),黄体期较长(约20天)。
2.激素反馈机制
卵泡期:雌激素对下丘脑产生负反馈,抑制GnRH脉冲的振幅→进而抑制FSH和LH释放。
围排卵期:雌激素转为正反馈,强烈刺激FSH和LH释放(尤其LH峰);此正反馈发生在下丘脑和垂体两个层面,形成“雌激素↑→GnRH/FSH/LH↑→更多雌激素↑”的循环,最终触发排卵。
3.狨猴的排卵特点
几乎每次排卵至少两个卵母细胞;多胎妊娠常见,但排卵未必同步;若发生胚胎死亡,可能最终表现为单胎妊娠。
4.黄体期调控
排卵后,雌激素迅速下降,孕酮显著上升。此时FSH/LH的脉冲频率被抑制(振幅不变),作用于下丘脑和垂体。抑制素B在卵泡早期浓度高。抑制素A在黄体期达峰值。黄体退化发生在月经前2–4天,随后新一轮卵泡发育启动。
5.卵泡发育与排卵机制
颗粒细胞不仅主导卵泡发育,还诱导周围基质细胞分化为膜细胞。参与卵泡发育的关键因子包括胰岛素样生长因子-1(IGF-1)、干细胞因子(SCF)、碱性成纤维细胞生长因子(bFGF)、TGF-β家族成员(如激活素、抑制素、骨形态发生蛋白BMPs、GDF9、TGF-β1/β2)、Hedgehog信号通路。
膜细胞在LH刺激下表达多种类固醇合成酶和转运蛋白:StAR(将胆固醇酯转运入线粒体)、P450scc(CYP11A)、P450c17(17α-羟化酶)、3β-HSD等。
排卵前,卵泡壁从产雌激素/雄激素转为产孕酮,防止闭锁并促进卵母细胞成熟。卵泡破裂依赖脂质、儿茶酚胺、血管活性物质及前列腺素。排卵后数小时内,颗粒细胞黄体化,大量分泌孕酮——此过程依赖LH。黄体期通过降低FSH活性和抑制素A分泌,抑制其他卵泡发育。若未妊娠,黄体持续14–16天,分泌孕酮、雌激素、抑制素A、松弛素、催产素。新世界猴(如狨猴)在黄体期的雌激素和孕酮水平显著高于人类和猕猴。受精发生在输卵管。尽管交配时有数百万精子进入生殖道,但在恒河猴中仅不到50个精子能到达输卵管。
(三)胎盘形成(Placentation)
1.食蟹猴作为人类胎盘模型
食蟹猴胎盘在绒毛结构、血-胎屏障、绒毛间隙血流等方面与人类高度相似。但存在关键差异:囊胚侵入子宫内膜较浅;形成双盘状胎盘(bidiscoid),而人类为单盘状(discoid);经历腔隙阶段(lacunar stage)后再建立母胎血液循环。
2.狨猴胎盘的独特性
初期形成小梁状附着(trabecular attachment);无腔隙阶段;血腔形成较晚,相比其他灵长类延迟。
3.人与猕猴胎盘的主要差异
猕猴滋养层壳边界更清晰;间质滋养细胞侵入较少;螺旋动脉侵入和母体循环建立更早;蜕膜反应模式不同。
4.免疫球蛋白(Ig)的跨胎盘转运
食蟹猴:研究检测了34种含Fc段的生物药。妊娠第100天后给药,胎儿药物浓度更高;但30/34种药物的胎儿浓度仍低于母体;IgG1转运效率最高(尽管IgG2/IgG4母体浓度更高)。
人类:胎儿IgG从孕13周起线性上升;足月时超过母体浓度;新生儿中IgG1和IgG4均高于母体。
乳汁与出生时Ig水平:乳汁中的Ig不怎么进入胎儿/新生儿血液循环;但乳汁IgA对肠道感染防护至关重要。
新世界猴(如狨猴):出生时IgG浓度不足成年水平的50%。
(四)妊娠期间的激素维持机制
在非妊娠状态下,垂体分泌的LH水平下降,导致黄体退化。而在妊娠期间,CG发挥类似LH的作用,维持卵巢黄体的功能,从而支持妊娠。整个孕期,母体自身产生的LH和FSH几乎可以忽略不计。以狨猴为例,妊娠第17天即可在母体血液和尿液中检测到CG。
此外,胎盘也参与孕酮的合成以维持妊娠。在卵泡中,膜细胞产生雄激素,由颗粒细胞将其转化为雌激素。而在胎儿体内,胎儿肾上腺(特别是其内侧占皮质4/5的“胎儿带”)产生脱氢表雄酮硫酸盐(DHEA-S),后者在胎盘合体滋养层细胞中被转化为雌二醇。雌二醇通过胎盘中的雌激素受体,促进合体滋养层的功能分化,尤其是诱导P450scc酶的表达——该酶是将胆固醇转化为孕酮所必需的关键酶。
在妊娠中期,母体皮质醇通过抑制胎儿垂体释放促肾上腺皮质激素(ACTH),抑制胎儿肾上腺皮质的发育。但到了妊娠晚期,胎盘中雌激素驱动的11β-羟基类固醇脱氢酶(11β-HSD)表达增加,将更多具有活性的皮质醇转化为无活性的可的松,从而解除对胎儿肾上腺的抑制,使其得以成熟并开始自主分泌皮质醇——这对胎儿肺部的成熟至关重要。
(五)胎儿生长调控
跨物种研究表明,胎盘是调控胎儿生长的核心器官。它能感知氧分压和营养状况,并通过分泌胰岛素样生长因子(IGFs)及其结合蛋白(IGF BPs)来调节胎儿生长——这两类因子作用相反。胎盘还表达大量转运蛋白,为胎儿提供营养支持。
在体型发育方面,猕猴和狨猴的生长数据已有报道。狨猴和绢毛猴(tamarins)具有一项独特特征:在妊娠第30天左右,多胎之间的胎盘循环会形成血管吻合,而绒毛腔则在妊娠第19至29天之间融合。值得注意的是,狨猴胎儿在妊娠前半期生长缓慢,直到妊娠中期后才加速生长,速度与其他灵长类相当。对妊娠115至158天的猴胎儿进行监测发现:动脉血氧饱和度51%–77%;血糖浓度平均29mg/dL(范围20–44mg/dL);动脉血压从妊娠115天的约35mmHg逐渐升至158天的50mmHg。
此外,狨猴(可能还包括绢毛猴)在灵长类中较为特殊:能在妊娠晚期吸收死亡胎儿而不发生死产;晚期胎儿死亡率较高,超声纵向监测显示,妊娠后半期胎儿死亡率达20–30%,而人类同期胎儿死亡率仅约10%。
(六)新生儿及幼年期
1.生理参数
Dawes等人报道了新生恒河猴的多项生理指标:动脉血氧饱和度约94%(胎儿期约为58%);血糖浓度约49mg/dL(胎儿期约为29mg/dL);血液携氧能力15–20mL氧气/100mL血液,从妊娠晚期到成年无显著变化;耗氧量受环境温度影响,在热中性区(能量消耗最低)约为35℃,新生恒河猴耗氧量约为14.8mL氧气/kg/min;出生时平均动脉血压55mmHg;体表面积出生时实测为0.122m²/kg,而成年恒河猴(体重5.56kg)体表面积为0.275m²,即0.049m²/kg;心率200–250次/分钟;心脏听诊通常无杂音,表明动脉导管已闭合。
此外,通过核磁共振代谢组学对血清和尿液的分析发现,恒河猴婴儿与人类婴儿在代谢物浓度上具有高度相似性:尿液中65%的代谢物浓度;血清中40%的代谢物浓度。二者相近,支持恒河猴作为研究人类婴儿发育的有效模型。
2.婴儿期激素变化
妊娠期间,胎儿肾上腺的“胎儿带”产生雄激素(如DHEA-S),由胎盘转化为雌激素。出生后,该区域发生退化。对于人类,胎儿带在出生后2周内迅速退化。恒河猴退化过程更缓慢,持续约6个月。在此期间,婴儿肾上腺活跃,DHEA、DHEA-S和雄烯二酮水平升高,峰值出现在约3.5月龄,此时网状带正在发育。
相比之下,狨猴胎儿带退化时间与人类相似,但不形成网状带。其皮质醇水平在出生后1周达高峰(约1900μg/dL),到2月龄骤降至<200μg/dL,并继续缓慢下降,至6月龄左右达到成年水平(约150μg/dL)。
3.出生后的生长发育
灵长类动物出生后的生长速率随年龄迅速下降。不同物种的出生体型如下:恒河猴坐高约195mm,体重约475g。食蟹猴出生体重约345g。狨猴出生体重仅约27g。
Gavan与Swindler通过坐高生长速率的对数转换,比较了恒河猴、黑猩猩和人类的生长模式:新生恒河猴初始生长速率为e1.263(每3个月增长3.5倍),略高于黑猩猩的e0.968(每3个月增长2.63倍)。两种动物的生长速率均快速下降,但恒河猴在5岁前始终维持更高生长速率。黑猩猩生长期比恒河猴更长,而人类生长期最长。因此,体型较大的灵长类并非长得更快,而是生长时间更长。
此外,对于营养正常母亲所生的恒河猴婴儿,出生时体型可预测其后续生长轨迹:体型较小的个体倾向于持续处于群体生长曲线的较低位置。
4.哺乳与早期营养
婴儿体重增长直接依赖于从母体乳汁中获取的营养量。灵长类乳汁的能量密度普遍低于其他哺乳动物:恒河猴乳汁平均能量密度约为1.0kcal/g(基于114只个体数据);狨猴乳汁约为0.76kcal/g。初产恒河猴母体泌乳量少于经产母体,但断奶时的体重和年龄主要取决于摄入的总乳汁能量。
乳汁不仅提供营养,还参与建立健康的肠道菌群。目前认为,携带特定有益菌的巨噬细胞可从肠道迁移至乳腺,并进入乳汁。在恒河猴乳汁中检测到五类需氧菌属:链球菌(Streptococcus)、肠球菌(Enterococcus)、乳杆菌(Lactobacillus)、片球菌(Pediococcus)和芽孢杆菌(Bacillus)。前三种也存在于人类母乳中。
哺乳是母体在子代发育过程中能量消耗最大的阶段。开始摄入固体食物的时间因物种而异:狨猴和食蟹猴约28日龄;恒河猴约46日龄。
所有灵长类均不能合成维生素C(缺乏L-古洛糖酸内酯氧化酶),必须从饮食中摄取。维生素D仅在缺乏紫外线B(UVB)照射时成为必需营养素:皮肤在UVB照射下可将7-脱氢胆固醇转化为维生素D3(胆钙化醇);维生素D3经酶促转化为活性形式1,25-二羟维生素D3(骨化三醇)。
两种维生素D补充形式:麦角钙化醇(D2,植物来源)、胆钙化醇(D3,动物来源)。新世界猴(如狨猴)对维生素D2利用效率极低,因此需通过UVB照射或直接补充D3,否则易患代谢性骨病。
5.性成熟发育特征
婴儿期、青春期和成年期的界定在文献中不统一,且个体差异显著。例如恒河猴的青春期激素启动时间范围为2.5–5.5岁。因此,相关年龄数据可作为参考,而非绝对标准。
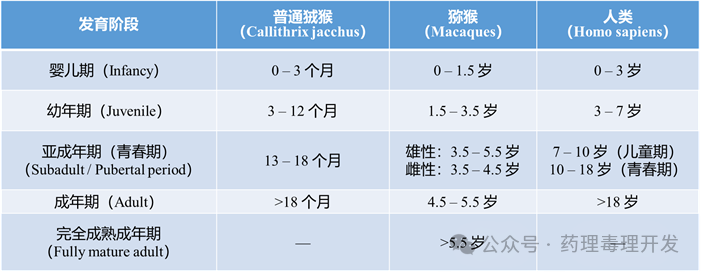
6.初潮(Menarche)时间
食蟹猴(印尼种群)初潮平均1.75岁,最晚4岁,50%个体在2.38岁前开始月经周期。恒河猴平均初潮年龄为2.25±0.28岁。尽管恒河猴与食蟹猴性成熟时间相近,但恒河猴为季节性繁殖(北半球繁殖期为10月至次年2月)。
7.婴儿期促性腺激素变化
雄性恒河猴:出生时FSH<400pg/mL,2周内升至500–1000pg/mL,维持约4周后,在1.5–8月龄逐渐下降。
雌性恒河猴:FSH在出生后首月升高,持续至3月龄,7月龄降至不可测水平。
人类婴儿也呈现类似模式,但FSH下降时间略早(3月龄后趋于稳定)。
LH变化:在男婴中上升更显著;去势猕猴和人类均显示LH与FSH趋势相似。狨猴新生儿FSH数据尚缺。
8.肾上腺功能激活
人类和猿类特有的现象:约6–7岁起肾上腺雄激素分泌增加(称为“肾上腺功能初现”),导致腋毛、阴毛生长及痤疮等第二性征。雄激素水平在20多岁达峰,之后缓慢下降。此过程独立于GnRH-垂体-性腺轴。猕猴和狨猴无肾上腺功能激活。
(七)性腺激活与青春期启动
青春期由GnRH脉冲释放恢复触发,标志着生殖系统“重启”。GnRH释放受身体状况(而非体型大小)调控,支持“体况感受器”(somatometer)假说。候选信号分子包括瘦素和IGF-1——高热量饮食可升高二者水平,与性早熟相关。在恒河猴中,Kiss1神经肽对GnRH释放至关重要。大脑整合性激素、IGF-1和季节性褪黑素信号以调控GnRH,机制仍不完全清楚。
雌性恒河猴青春期进程:20–30月龄:LH开始夜间脉冲式释放→引发初潮;随后20个月:LH水平持续上升,保留昼夜节律;50–60月龄:首次排卵,LH节律消失,全天均匀分泌。子宫随卵巢激素周期变化:雌激素促进子宫内膜增生并上调雌/孕激素受体;排卵后孕酮升高、雌激素下降;当孕酮>1ng/mL,受体表达下调,内膜对激素敏感性降低;若未妊娠,孕酮撤退→内膜脱落(月经)。
雄性恒河猴青春期进程:婴儿期LH升高(FSH轻度上升)→刺激睾丸间质细胞增殖并获得睾酮合成能力;LH刺激4天后,睾丸即出现脉冲式睾酮释放;此阶段Sertoli细胞增殖4倍,间质细胞增殖6倍。青少年期:Sertoli细胞静止,精原细胞持续增殖,间质细胞数量略减。从出生到成年,精原细胞数量增加约1000倍。青春期LH/FSH激增→Sertoli细胞达20亿/睾丸,间质细胞达1亿/睾丸;睾酮驱动精原细胞启动精子发生周期:生精上皮分4层,共12个形态阶段;完整周期(12阶段)耗时10.5天;从精原细胞到成熟精子共需42天。
1.生长突增(Growth Spurt)
雄性恒河猴:生长突增发生在睾丸成熟、睾酮升高之后,提示雄激素部分驱动;
雌性恒河猴:初潮比生长突增早约1.5年,此时仅达成年体重的30%;
人类女性:初潮时已达70%成年体型;
狨猴:推测生长突增发生于12–14月龄。
2.毒理学评估中的性成熟判定标准
雄性性成熟应以精液中检出精子为准,而非睾酮水平、睾丸大小、体重或年龄。雌性猕猴出现月经出血即可视为性成熟(尽管早期周期可能无排卵、不规律)。雌性狨猴无月经,性成熟需以排卵发生为标志,可通过尿液中孕二醇-3-葡萄糖醛酸浓度监测。通常≥18月龄视为性成熟,但群居社会结构可能显著延迟繁殖行为。
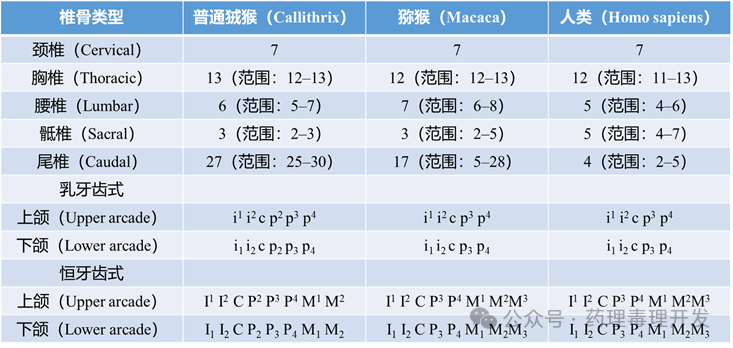
骨骼与牙齿发育成熟(Skeletal and Dental Maturation)Schultz对包括猕猴和人类在内的7–35种灵长类进行了详尽的生长发育比较研究,涵盖体重、身高及其他身体测量指标;骨化中心出现时间;骺板(生长板)闭合时序;颅腔容量;头骨、手足形态的精细绘图;牙齿萌出时间。
狨猴相关数据较为有限。Smith等人对33种灵长类的乳牙与恒牙萌出时间及模式进行了现代综述。最初研究动机之一是通过牙齿判断野生个体年龄,如今仍广泛用于评估化合物对骨或牙发育的影响。狨猴、猕猴和人类椎骨数量和牙齿发育对比如下表所示。
心血管生理学对比
(一)心脏解剖结构
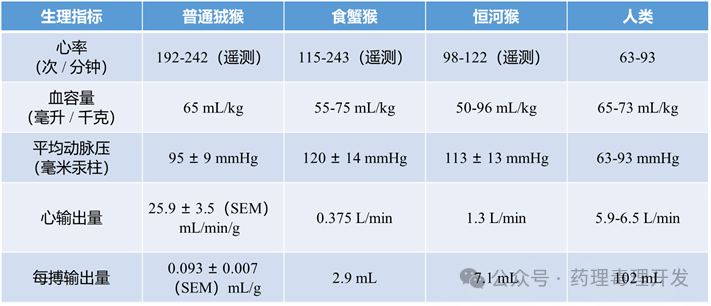
不同种属心血管参数如下表所示:
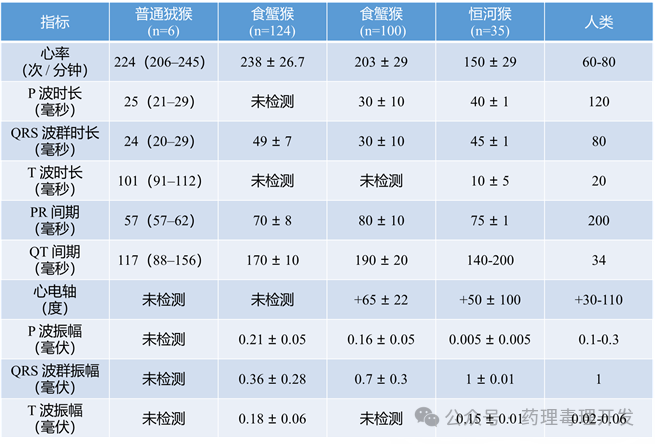
不同种属电生理参数如下表所示:
猴与人类的心血管系统总体上非常相似。
恒河猴:Swindle等人对28只雌性和19只雄性恒河猴(年龄从胎死到>24岁)的心脏进行了形态测量研究。结果显示,心脏各部分的比例在整个生命周期中保持高度一致;唯一的性别差异是雄性的二尖瓣/三尖瓣环周长比大于雌性。
狨猴:其心脏重量占体重的比例为0.51±0.38%,与其他哺乳动物一致,且无显著性别差异。
颈动脉与Willis环:与人类相比仅有细微差异,但可能影响脑血管梗死模型的构建。
自主神经支配:灵长类与人类在迷走神经支配方面无差异;交感神经节位置略偏头端,类人猿具有更明确的胸心神经。
发育变化:新生恒河猴的心率主要受交感神经调控,而成年后则由副交感神经主导——这对药物干预自主神经系统的效果具有重要意义。
(二)心肌收缩调控
心肌细胞兴奋-收缩耦联及心率的自主调控涉及多种离子通道,相关机制已有综述。与人类相比,啮齿类动物(如大鼠、小鼠)在动作电位时程、肌球蛋白重链亚型及结构蛋白异构体表达方面存在显著差异,这些差异反映了物种对各自生理需求的适应。
不同物种对药物或毒性物质引起的离子通道干扰敏感性不同。例如,在预测致心律失常风险方面,非人灵长类有时优于犬类。
尽管自主神经调控在发育过程中发生变化,但在成年个体中,呼吸对心率的影响(即呼吸性窦性心律不齐)在多个物种中普遍存在。
(三)尖端扭转型室性心动过速
定义与特征:TdP是一种多形性室性心动过速,心电图表现为QRS波群振幅呈正弦样波动,形似“扭曲的彩带”。其发生常与QT间期延长相关。
药物安全性评估:目前通过标准化测量药物对QT间期的影响,并计算校正QT间期(QTc)来评估风险。需注意,健康狨猴的QTc存在昼夜节律波动。
适用动物模型:犬、食蟹猴、小型猪、狨猴、豚鼠和人类均可用于可靠检测药物对QT间期的影响;而大鼠和小鼠因心脏离子通道表达与功能差异较大,不适合作为TdP模型。
主要靶点:HERG通道(编码快速延迟整流钾电流IKr)是药物诱导QT延长最常见的作用靶点。典型药物如多非利特。已有食蟹猴发生TdP的个案报道。
模型选择考量:尽管犬仍是心血管安全性评价的主要体内模型,但在生物制剂代谢、靶点结合特性等方面存在种属差异时,非人灵长类可能更为必要。
(四)TdP的潜在机制
HERG通道抑制导致TdP的确切机制尚不完全清楚,但可能与心肌细胞在不同区域的离子电导特性差异有关,尤其在肾上腺素能刺激下更为明显。中层心室肌细胞的动作电位更容易延长,可能成为触发TdP特征性室性早搏的起源点。有假说认为,只有那些导致心外膜与中层心肌细胞去极化特性差异增大的药物才真正具有诱发TdP的风险。这种区域电活动差异的分子基础可能与缝隙连接蛋白Connexin 43的空间表达差异有关。
人类存在性别对自主神经刺激的心肌反应差异,这一现象在恒河猴中也得到了复现。
(五)心电图(Electrocardiography)
遥测心电图技术在药物安全性评价中已与呼吸生理监测和自动化采血系统联用,显著提高了对心律失常风险的检测敏感性。
一项对长期植入遥测装置的食蟹猴的研究显示:白天最常见的心律失常包括:房性早搏(10.7%)、逸搏(3.8%)、窦房结停搏(2.8%)和室性早搏(VPB,25.4%);夜间则以窦房结停搏(32.6%)、逸搏(18.5%)和VPB(11.7%)为主,表现明显不同。相比之下,人类24小时内至少出现一次VPB的比例约为39%,但前瞻性研究发现这类心律失常并不能预测未来冠状动脉事件。建议在正式实验前对个体动物进行基线监测,以控制自发性心律失常频率的个体差异。
在化学镇静状态下,对黑耳狨猴的心电参数研究显示其正常值范围无性别差异。恒河猴的心电激动时间(61毫秒)显著短于人类(97毫秒),该差异不能完全由静息心率不同解释,提示恒河猴心肌细胞可能具有更高的兴奋传导速度。除激动时间外,年龄和性别对其他心电参数的影响与人类相似。
肾脏生理学对比
关于非人灵长类肾脏基础生理的研究文献在1970年代达到高峰。目前人类肾脏生理的基础知识已非常完善,且在非人灵长类中具有良好的可转化性。下表汇总了狨猴、猕猴和人类的部分基本肾脏生理参数。
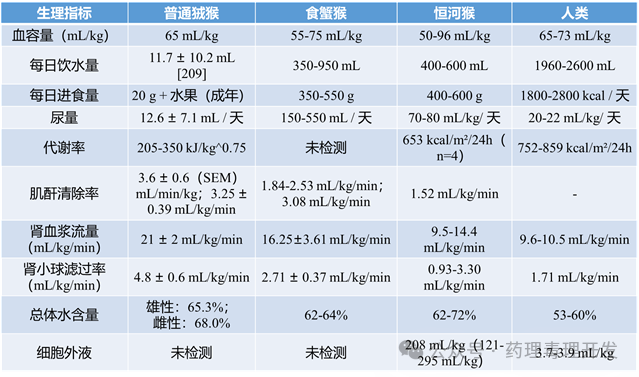
(一)肾小球滤过率(GFR)与肾血流
在一项针对6只未成熟和7只成年恒河猴(年龄和性别未说明)的研究中:菊粉清除率(inulin clearance,GFR金标准)为:未成熟猴:2.18±0.680mL/min/kg;成年猴:1.96±0.445mL/min/kg。作为对比,人类女性的菊粉清除率为1.75±0.22mL/min/kg。同一研究中还测得:肌酐清除率:3.08±0.939mL/min/kg→表明恒河猴肾小管对肌酐存在分泌作用(即肌酐不仅被滤过,还被主动排泌)。对氨基马尿酸(PAH):8.06±1.220mL/min/kg(用于估算肾血浆流量)。
Liu的研究报道了成年雌性恒河猴的多项肾功能参数(按体表面积标准化后更接近人类值):总肾血浆流量(RPF):557±153mL/min/m²;总肾血流量(RBF):809±217mL/min/m²;肾氧耗量:1010±433mL/hr/m²;;葡萄糖最大重吸收速率(TmG):67.4±14.3mg/min/m²。Liu与Higbee还通过直接测量猕猴体表面积,建立了基于头臀长或头足长的最佳体表面积估算公式,有助于跨物种生理参数的标准化比较。
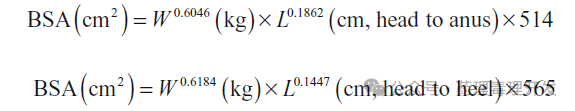
(二)水摄入与尿液排泄节律
对20只恒河猴的观察显示:排尿高峰出现在早晨7–8点,约在喂食前1.5小时;饮水主要集中在下午早些时候;性别间无显著差异。平均24小时数据。饮水量:83±31mL/kg;尿量:51±19mL/kg;成年恒河猴的肾脏重量和体积高度一致,即使未按体重校正也变异较小:雄性肾脏比雌性重约0.8g;左肾比右肾重约0.5g;雄性左肾平均重13.7g,体积13.5mL。
呼吸生理学对比
一、解剖结构
鼻腔
恒河猴鼻腔中没有类似人类的上鼻甲;其上鼻甲相当于人类的中鼻甲。存在上颌窦,但无蝶窦。鼻腔黏膜类型包括鳞状、移行、呼吸性、嗅觉性和淋巴上皮黏膜。与人类相似,恒河猴的嗅觉上皮面积显著退化。恒河猴鼻腔容积约为8mL,而人类为16–28mL。雌性恒河猴嗅觉黏膜中的肥大细胞数量在排卵期达到高峰;雄性则维持中等水平。电镜观察显示,雌猴嗅觉上皮中的支持细胞在排卵周期中有形态变化,可能与性激素调控的嗅觉功能相关。
2.犁鼻器
狨猴保留犁鼻器(尽管比原猴亚目小),而恒河猴和人类均无。
3.喉部
恒河猴喉部存在一个喉气囊,位于甲状软骨与舌骨之间,开口于会厌尾侧、略高于室带处。曾认为其作为共鸣腔,但手术切除后未改变发声频率,功能尚不明确。
4.肺叶结构
恒河猴左肺有3叶(人类为2叶);右肺有4叶:头叶、中叶、尾叶和副叶(人类无副叶)。
5.气道上皮细胞
成年恒河猴气道上皮含4种细胞:纤毛细胞、杯状细胞、小黏液颗粒细胞、基底细胞。Clara细胞(非纤毛细支气管上皮细胞)仅见于呼吸性细支气管。恒河猴Clara细胞的超微结构(顶质富含膜包颗粒、粗面内质网丰富、无滑面内质网)与人类相似。
6.狨猴肺结构数据
成年狨猴:肺泡表面积7662±1647cm²;毛细血管表面积6000±1549cm²;肺毛细血管容积1.01±0.34mL;氧扩散能力0.0299±0.0134mL O₂/sec/mmHg(相对于大型啮齿类属较高水平)。
二、发育过程
气管发育
<46天胎龄仅单层柱状非纤毛细胞。46天起出现纤毛细胞。120天,四种主要细胞类型均已出现。134天,气管上皮结构接近成体。
气管腺体:80天开始形成黏膜下细胞柱,先分化黏液细胞,后为浆液细胞,持续至出生后。
2.喉部生长
对1–51日龄恒河猴及食蟹猴的研究显示,喉部尺寸个体差异大,与日龄无明显相关性。
3.肺发育(恒河猴)
50–75天原始细支气管增生。100天首次出现肺泡。150天肺泡占肺容积50%(成体为2/3);I型和II型肺泡细胞已可区分。
磷脂成熟:约162天时,磷脂酰胆碱上升,磷脂酰乙醇胺和鞘磷脂下降,同时糖原减少。尽管人类肺成熟受皮质醇调控,但在恒河猴中未发现皮质醇与上述指标的明确关联。
4.Clara细胞分泌蛋白(CCSP)
出生后1个月含量低,3个月达近成体水平,提示1–3月龄是气道关键发育期。
5.表皮生长因子(EGF)作用
给胎猴注射EGF可加速II型肺泡细胞分化,并增加表面活性蛋白A,但不影响整体肺生长。
6.出生后肺泡数量
对2–13岁恒河猴的立体学研究显示:肺体积增长源于现有肺泡扩张,无新肺泡生成。
三、呼吸生理
基础参数
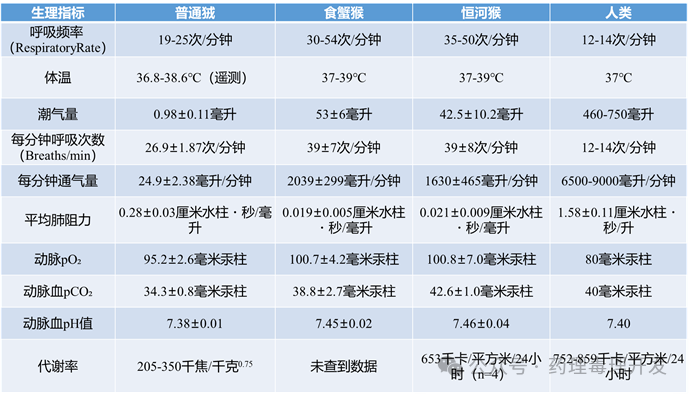
恒河猴、食蟹猴和狒狒的多项呼吸生理参数已被测定并相互比较。
恒河猴的最大呼气流速与年轻成人人类相当。胸壁顺应性方面,恒河猴和冠毛猴的胸壁比狗僵硬约4倍;肺顺应性约为犬的2倍;导致功能残气量(FRC)占比高达64%(约为犬的2倍)。
2.哮喘模型与药理反应
组胺气雾剂可使气道阻力增加200%,肺顺应性下降30–80%。恒河猴与食蟹猴对多种支气管扩张剂(如前列腺素、特布他林、异丙肾上腺素、氨茶碱)反应一致,表明二者肺生理高度相似。
离体肺切片实验比较不同灵长类对收缩剂的反应:组胺:在猕猴中作用最强,狨猴仅为其一半;白三烯D4:狨猴和恒河猴无反应,但食蟹猴和狒猴出现50%气道收缩,更接近人类。说明,食蟹猴比恒河猴更适合作为人类支气管收缩反应的预测模型。
3.颗粒物沉积模型
Asgharian等建立几何模型预测:婴儿及青年恒河猴对10μm颗粒的肺部沉积率低于人类。其他模型也证实猴子鼻腔截留率更高,肺部沉积率更低,与人类存在种属差异。
神经生物学对比
(一)行为与认知的相似性与差异
非人灵长类(如恒河猴、食蟹猴、绿猴)的大脑发育、解剖结构和功能与人类高度相似,因此被广泛用作神经科学研究模型。一些特定行为也表现出跨物种一致性。例如:猕猴和绿猴中存在性别相关的玩具偏好(雄性偏好车辆类,雌性偏好玩偶类),与人类儿童的发现一致。已建立针对猕猴的神经行为评估测试体系,用于检测药物对神经系统的不良影响,其设计类似于啮齿类动物的测试方法。
在认知能力比较中:黑猩猩在空间理解与工具使用方面优于食蟹猴;但在社会认知任务(如合作、意图理解)中并无优势;人类儿童则在社会认知领域显著领先。这些功能差异背后必然存在分子与解剖基础,但目前理解仍处于初级阶段。
(二)人类与非人灵长类的分子差异
Geschwind与Rakic最近综述了驱动认知差异的关键分子机制,包括:超过600个基因在人类谱系中经历了加速进化;500多个高度保守区域在人类中发生缺失;400多个区域存在组蛋白甲基化差异;至少8个基因家族在人类中稳定扩增;24个基因间区在人类中高转录,可能具调控功能;至少10种新microRNA为人类特有;300多个增强子区域特异性活跃于人类皮层;54个基因在前额叶皮层中人类特异性上调。
代表性基因举例:1)FOXP2:人类版本有两个氨基酸突变;影响纹状体环路,改变感觉运动整合;可能还参与颅面发育和神经前体细胞增殖;2)SRGAP2C:是人类特有的SRGAP2A截短形式;在转基因小鼠中可增加树突棘密度、延长棘颈,使其更接近人类神经元形态。
此外,人类额叶中共表达基因网络比黑猩猩和猕猴更复杂,尤其在调控神经元形态和突触功能方面。
综上,大量分子层面的差异可能是人类高级认知能力的生物学基础。
(三)比较神经解剖学与发育机制
皮层发育的“径向单元假说”
皮层生长分为两阶段:脑室区神经干细胞增殖→决定皮层表面积;神经前体细胞增殖与径向迁移→形成新皮层各层结构。
神经发生持续时间显著不同:小鼠约6天;猕猴约60天(胚胎第40–100天);人类约100天(胚胎第43–120天);对应新皮层表面积比例约为1:100:1000。
2.区域特异性进化
例如:灵长类初级视皮层第IV层神经元厚度比其他哺乳动物增加4倍(该层接收传入信号);这种变化可能由神经发生第二阶段中区域特异性的分子调控所驱动。
3.基因表达梯度差异
产前发育期间,人类与猕猴在新皮层基因表达梯度上存在差异;这些差异可能指导人类特有脑区(如布罗卡区、韦尼克区等语言中枢)的形成。
4.青春期后的基因表达分化
青春期后,人类与猕猴的基因表达再次显著分化;人类表现出更高复杂性,可能支持高级认知功能的成熟。
(四)突触发育与修剪
在皮层构建完成后,神经元之间开始大量形成连接(突触发生)。
灵长类特有模式:突触密度在儿童期达峰值(2岁人类的突触数量约为成人的2倍);高密度状态维持较长时间;到性成熟前后经历大规模突触修剪。
Watson等人对大鼠、猕猴和人类的出生后脑发育进行了系统比较,强调:理解各物种发育事件的时间对应关系,对于非临床安全性评价至关重要;可据此选择合适的动物模型和给药时间窗,确保药物暴露覆盖可比的发育阶段。
视觉系统对比
(一)人类视觉系统的两大特化特征
三色视觉和中央凹。中央凹是视网膜上一个直径约0.5mm的凹陷区域,具有以下特点:仅含视锥细胞(无视杆细胞);排除其他细胞体和血管(无血管区);光感受器→双极细胞→神经节细胞之间形成一对一连接通路,实现最高视觉敏锐度。中央凹并非人类独有:除原猴亚目和夜行性的猫头鹰猴外,所有灵长类均具备中央凹。其他动物也有类似高分辨率结构,如:猛禽(如隼):拥有两个中央凹(分别用于双眼或单眼高分辨视觉);其他哺乳动物:具有视觉条带或中央区。
(二)猕猴作为视觉研究的主要模型
猕猴眼球在光感受器密度和光谱敏感性方面与人类高度相似,是视觉研究的主流模型。中央凹视锥细胞密度对比:恒河猴140,500个/mm²;食蟹猴100,400个/mm²;人类平均199,000个/mm²(范围:98,000–324,000个/mm²)新世界猴(如狨猴):不同物种间中央凹视锥密度相近;中央凹直径不随体型变化;狨猴中央凹密度为152,000–211,000个/mm²。由于中央凹无血管,依赖扩散供氧和营养,其直径受扩散效率限制。因此,眼越大,中央凹所占视角越小。狨猴约7.8°,人类<5°。
(三)色觉与视蛋白分子遗传学
光谱敏感性
猕猴视锥细胞最大敏感波长:短波(蓝)430nm;中波(绿)530nm;长波(红)561nm;视杆细胞491nm。人类与食蟹猴的红、绿视锥光谱敏感性几乎完全一致。
2.视蛋白基因复杂性
短波视蛋白基因(S-opsin)位于常染色体;中波(M)和长波(L)视蛋白基因:由X染色体上的基因重复产生;基因间重组或突变可导致多种视蛋白表达组合。
3.色盲发生率差异
人类:约1%的高加索男性患红绿色盲(二色视);食蟹猴:红绿色盲极为罕见。
4.狨猴的特殊色觉机制
狨猴的视杆视蛋白基因也位于X染色体(另有一个常染色体短波视蛋白基因);X染色体上存在三个等位基因,最大敏感波长分别为543nm、556nm、563nm。雄性狨猴(仅一条X染色体):总是二色视;雌性狨猴(两条X染色体):若携带不同等位基因,通过X染色体随机失活(X-inactivation),可在不同视锥细胞中分别表达两种L/M视蛋白,从而实现三色视觉。



-
 大小鼠移动式超净/微屏障饲养系统
大小鼠移动式超净/微屏障饲养系统 -
 大小鼠独立送风IVC饲养系统
大小鼠独立送风IVC饲养系统 -
 大小鼠代谢笼
大小鼠代谢笼 -
 玩具葫芦
玩具葫芦 -
 大小鼠连续耳标钳
大小鼠连续耳标钳 -
 不锈钢猴镜
不锈钢猴镜 -
 猴玩具球
猴玩具球 -
 猴项圈
猴项圈 -
 捕猴杆
捕猴杆 -
 捕猴网
捕猴网 -
 舒泰50(高纯试剂,仅供试验用)
舒泰50(高纯试剂,仅供试验用) -
异氟烷(调试麻醉机试剂)
-
 鼠断头器
鼠断头器 -
 摇铃
摇铃


400 - 102 - 1174






咨询热线

莱
艾
特
商
城


扫
一
扫
关
注

京ICP备14028330号-1 京公网安备 11010802039490号
Copyright © 2015 All Rights Reserved www.bjlat.com 设计维护:北京莱艾特科技发展有限公司
郑重声明:引用他人的信息,本网站均已经列明出处。
转载的目的在于学术交流与讨论,若有任何侵犯您的权益之处,请与我们联系删除相关内容。
地址:北京市海淀区天秀路10号中国农大国际创业园3号楼2层2002 邮箱:info@bjlat.com 电话:010-62969989;13661392264